Разработанное оборудование,включая специальное программное обеспечение,
прошло испытания на биополигоне Агрофизического института Санкт-Петербурга.
Примеры использования фитомониторов
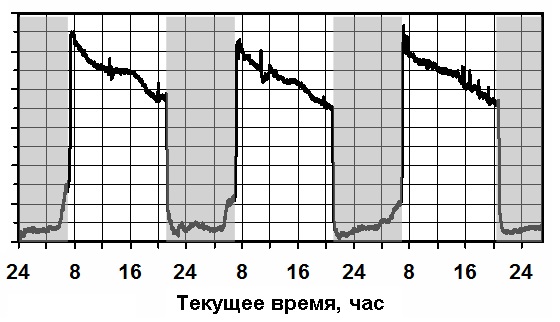 Рис.1
Рис.1
В комплект датчиков устройства "Phytoscan-3T" входит датчик интенсивности
сокодвижения от корневой системы растения к испаряющим поверхностям листовых
пластинок. По результатам регистрации этого параметра (Рис.1) виден циклический
характер с суточным циклом. Включение света приводит к началу транспирации.
В свою очередь интенсивное испарение с поверхности листа вызывает дефицит
влаги в листе. Это является причиной непрерывного
движения жидкости по сосудистой системе растения от корня к листовым поверхностям.
На рисунке показан характерный вид такого движения в течение световых периодов.
Выключение света приводит к прекращению
транспирации и, сответственно, резкому снижению скорости перемещения
жидкой составляющей по сосудистой системе.
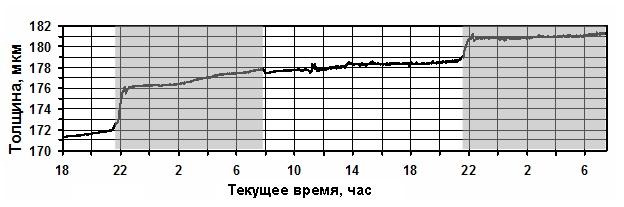 Рис.2
Рис.2
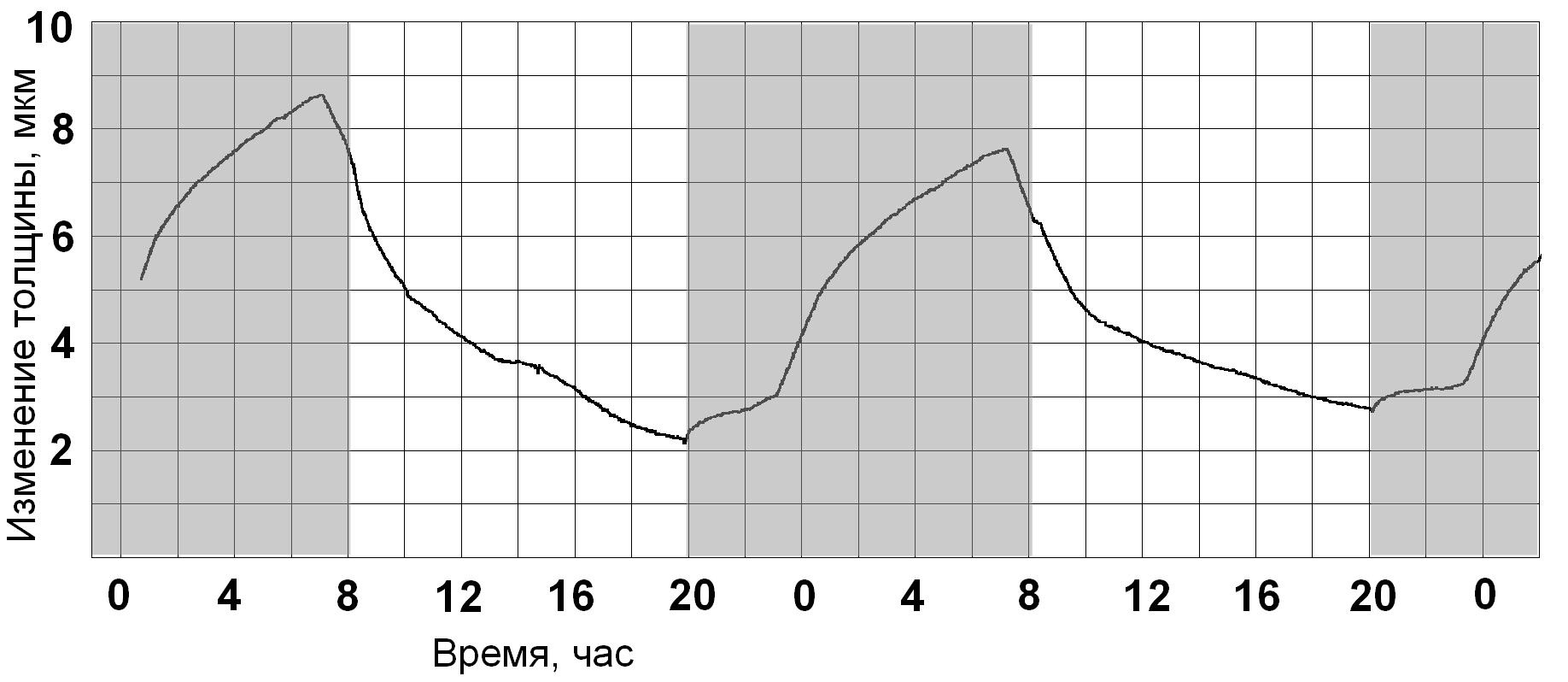
Если на листе установить датчик микроизменений его толщины, то в условиях оптимизации
микроклимата график будет иметь положительный тренд (Рис.2). Он характеризуется непрерывным ростом
толщины листовой пластинки, как в ночной, так и в дневной периоды.
Характерно то, что ночной прирост всегда выше, чем дневной.
Подобные данные позволяют
осуществлять оптимизацию микроклимата, не только за счёт корневого и
газового питания, но и путём рациональной организации световой среды (интенсивность,
фотопериод, спектральный состав).
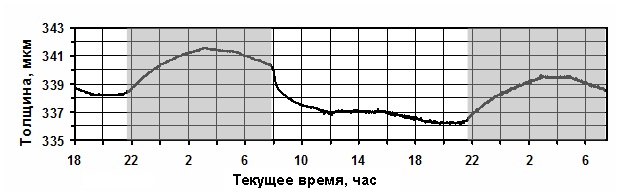 Рис.3
Рис.3
В то же время, можно увидеть, что с помощью нашего устройства можно очень точно
установить величину дефицита водного питания и довольно точно момент начала
дефицита влагообеспечения. На рисунке можно увидеть (Рис.3), что толщина листьев при
недостатке полива может даже уменьшаться, т.е. график имеет отрицательный тренд.
Степень уменьшения оводнённости можно увидеть путём сравнения текущей толщины
листовой пластины с таковой в предыдущие сутки.
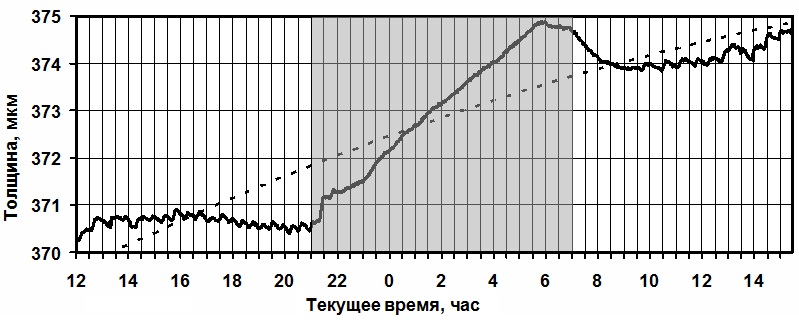 Рис.4
Рис.4
Можно наблюдать, что когда нижние листья испытывают дефицит влагообеспечения,
верхние ярусы, находящиеся под интенсивным световым облучением, продолжают
суточный прирост толщины листовой пластины, что видно на данном графике (Рис.4).
Подобные явления заставили увеличить количество параллельных каналов в
устройствах фитомониторинга, что реализовано в Physcan-4T.
На разных частях контрольного растения (на различных ярусах) устанавливаются
подобные датчики.
Специальное программное обеспечение обобщает параллельные сигналы,
которые являются входными параметрами для виртуального образа растения
в виде системы дифференциальных уравнений.
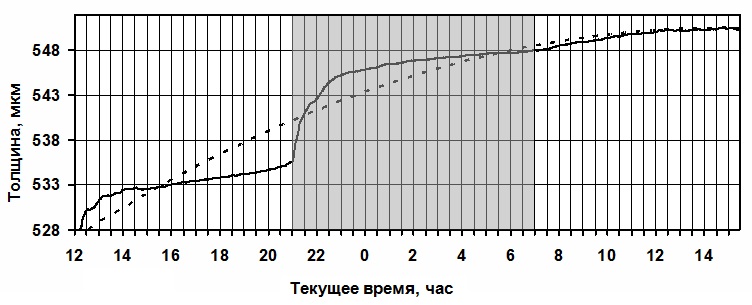 Рис.5
Рис.5
Существует оптимальная величина освещённости, изменение её в сторону
увеличения или уменьшения приводит к снижению интенсивности суточного прироста
(уменьшению угла положительного тренда).
Например, на графике (Рис.5) второй световой период характеризуется
несколько меньшей интенсивностью. Что приводит к уменьшению скачка
при включении света и снижению общей величины суточного прироста.
Аналогичная картина наблюдается при увеличении интенсивности за пределы оптимума.
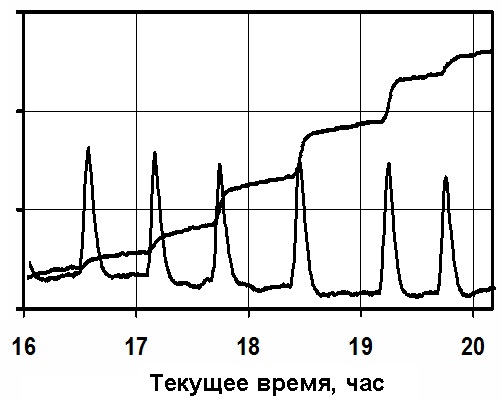 Рис.6
Рис.6
На рисунке 6 график восстановления оводнённости листа после стресса
(недостаток полива или дефицит влажности воздуха).
Датчик скорости ксилемного потока от корневой системы к листьям показывает,
что после падения
интенсивности сокодвижения при недостатке полива его восстановление происходит
прерывистыми перемещениями жидкости - нижняя линия на рисунке. Следствием этого
является ступенчатое восстановление оводнённости листа, что показано на верхней
линии рисунка.
Этот процесс связан со структурными изменениями проводящей
системы растений - увеличивается её сопротивление.
В результате, даже кратковременный недостаток влаги в объёме листовой пластинки
с последующим, неполным востановлением тургора не проходит бесследно.
Замедление роста после водного стресса неминуемо снижает растительную массу и
замедляет развитие плода по сравнению с растениями находившимися в оптимальных
условиях развития.
Динамическая реакция
Стевии (STEVIA REBAUDIANA BERTONI L.) на изменение фотопериода и влажности
почвы.
Карманов И.В., Кочетов А.А.
(в сокращении 2014)
Серия опытов по приспособляемости не только к дефициту влаги, но и к
изменениям интенсивности освещённости установила границы допустимого климатического
стресса. Свет и фотопериодический режим выращивания являются главными регуляторами
суточного хода всех физиологических процессов в растении. Характер изменений параметров
водного обмена в условиях оптимального водообеспечения обусловлен преимущественно
сменой дня и ночи, то есть изменением интенсивности транспирации. В качестве основного
объекта наших исследований была выбрана стевия Stevia rebaudiana Bertoni L. -
новая культура из Южной Америки, в листьях которой содержится комплекс сладких
веществ - дитерпеновых гликозидов, являющихся диетическими сахарозаменителями.
Растения стевии требовательны к влагообеспечению почвы и к освещённости.
Они приспособлены к непрерывному потреблению влаги во время вегетации.
Анализ суточной динамики тургорного напряжения листовой пластинки показывает,
что на опережающем уровне происходит подстройка к будущим предполагаемым циклическим
изменениям освещённости. Сдвиг фаз сохраняет своё значение по отношению к времени
включения/выключения света независимо от тренда общего структурного прироста толщины
листа.
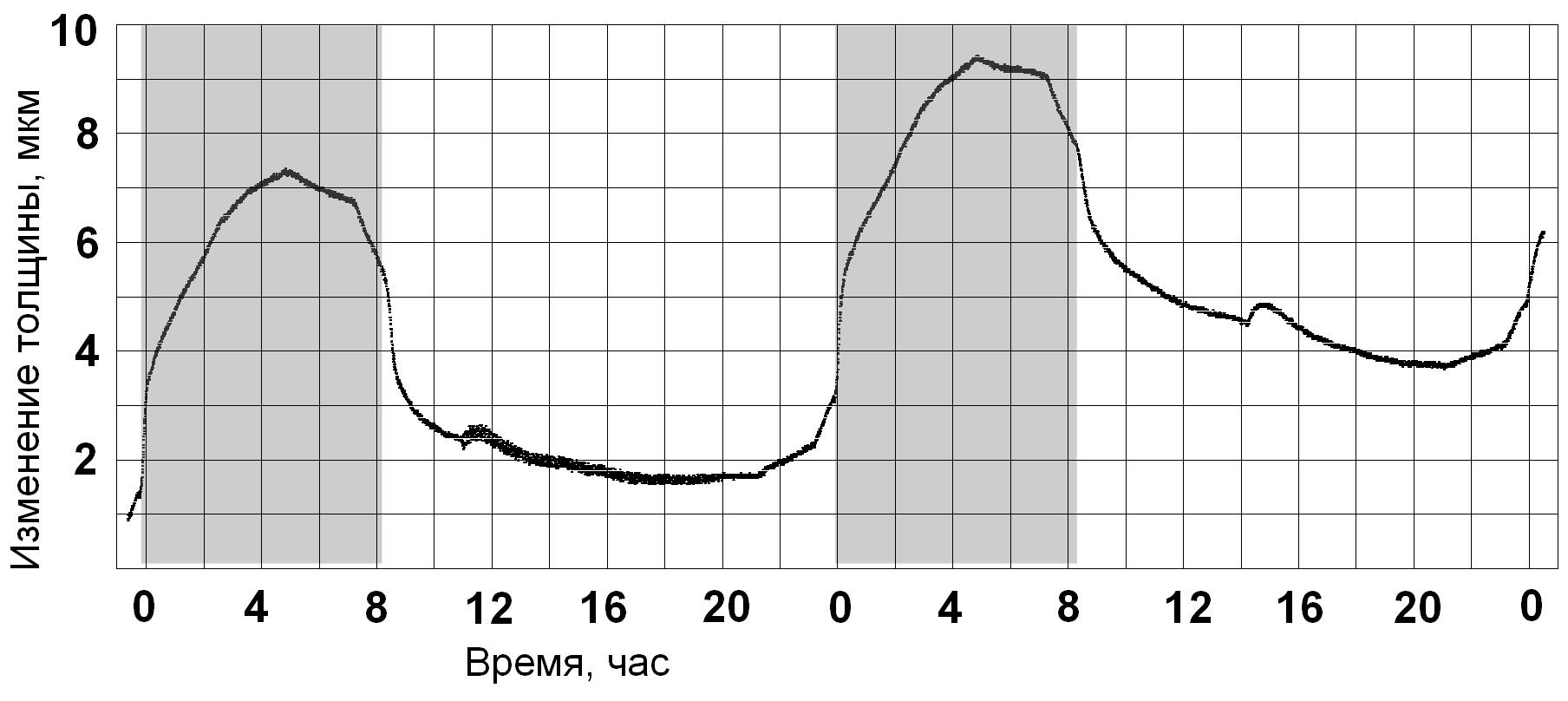
Cуточная динамика изменения толщины растущего листа у растения, которое
выращено при 16- часовом фотопериоде.
- включение света вызывает снижение тургора с постепенной стабилизацией и
некоторым повышением в конце светового периода;
- момент полива отражается последующим непродолжительным увеличением тургора
с дальнейшей стабилизацией (на графике - полив в первые сутки в 11 часов,
во вторые - около 14 часов);
- выключение света вызывает плавное повышение тургора, связанное со
снижением транспирации в темновой период и, следовательно, с возрастанием
оводнённости листа;
- вторая половина темнового периода характеризуется почти линейным уровнем
тургора с незначительным его снижением в конце;
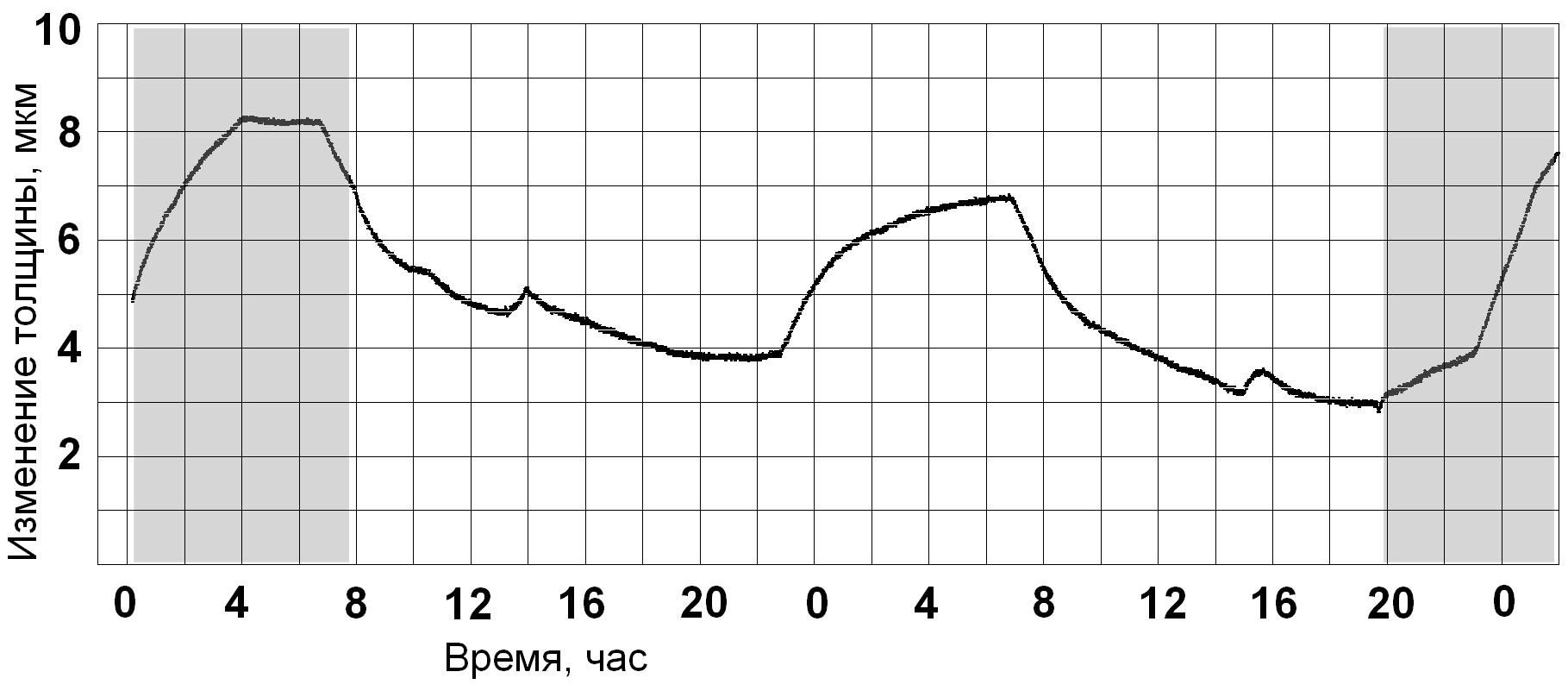
Устойчивость суточной динамики изменения толщины листа при 16-часовом
фотопериоде подтверждается её видом при переводе растения в условия
непрерывного освещения. В первые сутки в дневное время характер кривой
сохраняется, но затем в условиях непрерывного освещения наблюдается ярко
выраженная реакция на мнимое выключение света в виде излома кривой с "ночным"
возрастанием с последующим изломом кривой в сторону убывания в "обычное" время
перед "включением" света. Несмотря на сохранение объемов, и времени поливов,
наблюдается отрицательный тренд и снижение максимального значения тургора
в "ночное время".
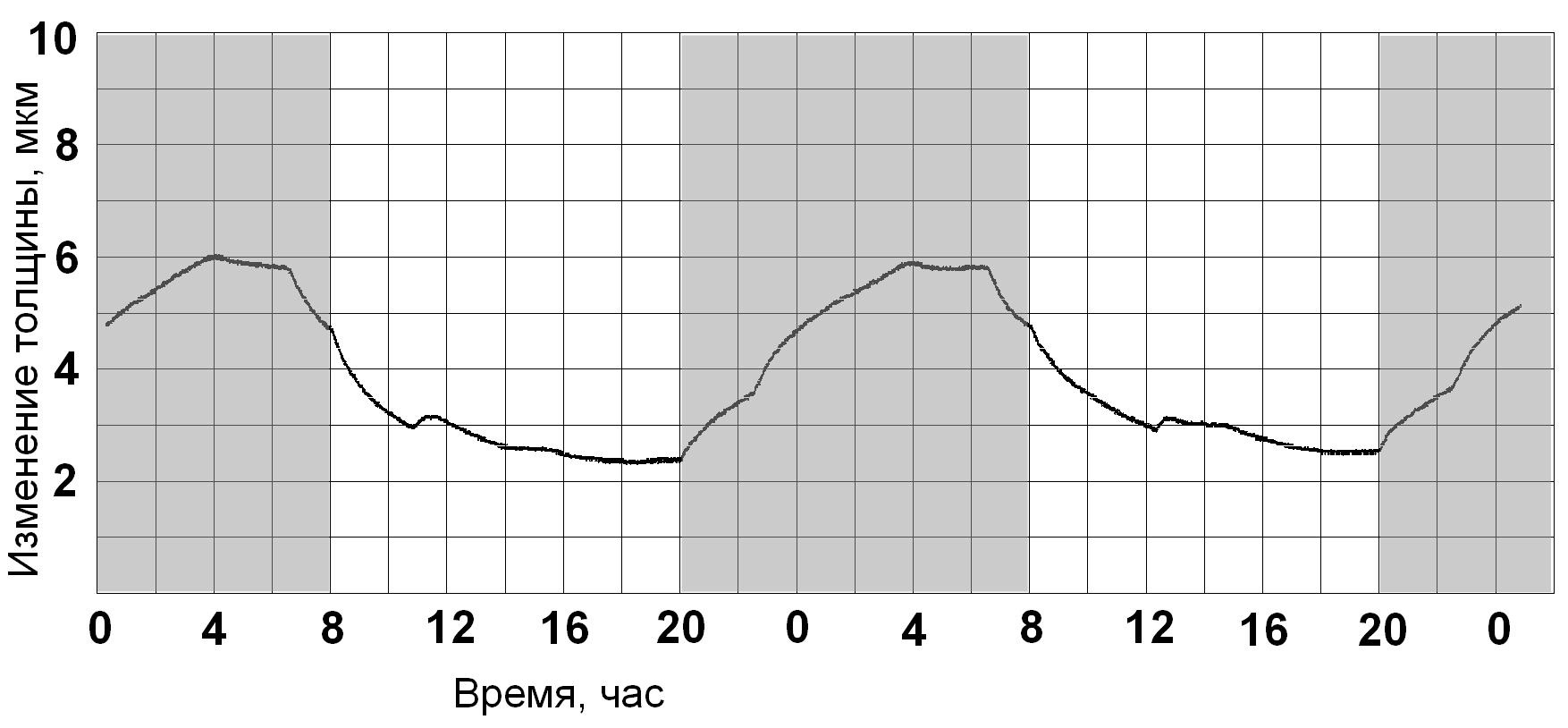
Перевод с 16-часового фотопериода на 12-часовой. При этом выключение света
осуществляся на 4 часа раньше по сравнению с привычным для
растения 16-часовым фотопериодом.
Изменения в динамике кривой тургорного напряжения:
- нарушение монотонности циклической зависимости тургора листа от времени;
- отсутствие периода стабилизации между возрастанием тургора и дальнейшим
его снижением в конце темнового периода;
- появление явно выраженной реакции на "неожиданное" выключение
света (в 8 часов вечера) и переход к равномерному возрастанию тургора
при наступлении полуночи (т.е. времени, обычного для растения выключения света).
Через 10-15 суток можно наблюдать физиологическую приспособляемость
растения к новому световому циклу. Кривая зависимости тургора листа от
времени приобретает более плавный монотонный вид.
